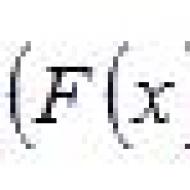Характеристика основных экологических групп растений. Строение листа растения, типы расположения листовых пластин, фотосинтез и транспирация Растения с большим количеством устьиц примеры
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток, между которыми имеется своеобразный межклетник, или устьичная щель ( рис. 37). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью располагается дыхательная, или воздушная, полость, окруженная клетками мякоти листа . Клетки эпидермы, примыкающие к замыкающим, получили название побочных, или околоустьичных. Они участвуют в движении замыкающих клеток. Замыкающие и побочные клетки образуют устьичный аппарат. Число побочных клеток и их расположение относительно устьичной щели позволяют выделять ряд устьичных типов. Их изучением занимается стоматография. Данные стоматографии нередко используются в систематике растений для уточнения систематического положения таксонов. Наиболее часто встречающиеся устьичные типы показаны на рисунке 38 .
Аномоцитный тип устьичного аппарата обычен для всех групп высших растений , исключая хвойные . Побочные клетки в этом случае не отличаются от остальных клеток эпидермы. Диацитный тип характеризуется только двумя побочными клетками, общая стенка которых перпендикулярна устьичной щели. Этот тип обнаружен у некоторых цветковых , в частности у большинства губоцветных и гвоздичных . При парацитном типе побочные клетки располагаются параллельно замыкающим и устьичной щели. Он найден у папоротников , хвощей и ряда цветковых растений . Анизоцитный тип обнаружен только у цветковых растений . Здесь замыкающие клетки окружены тремя побочными, одна из которых заметно крупнее или мельче остальных. Тетрацитным типом устьичного аппарата характеризуются преимущественно однодольные . При энциклоцитном типе побочные клетки образуют узкое кольцо вокруг замыкающих клеток. Подобная структура найдена у папоротников , голосеменных и ряда цветковых . Расположение замыкающих клеток относительно прочих клеток эпидермы у разных видов неодинаково. В одних случаях замыкающие клетки находятся на одном уровне с эпидермальными, иногда выступают над ними или, напротив, залегают значительно глубже (погруженные устьица). Последнее наблюдается у растений, приспособленных к засушливым условиям. Иногда углубления, в которых располагаются устьица, выстланы или прикрыты волосками. Называются они устьичными криптами .
Число и распределение устьиц на листе или побеге варьируют в зависимости от вида растений и условий жизни. Число их обычно колеблется от нескольких десятков до нескольких сотен на 1 кв.мм поверхности.
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при недостаточном водоснабжении в ночные часы, а иногда и днем тургор в замыкающих клетках понижается и щель замыкается, снижая тем самым уровень транспирации. С повышением тургора устьица открываются. Считают, что главная роль в этих изменениях принадлежит ионам калия. Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов . Первичный крахмал хлоропластов , превращаясь в сахар , повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и переходу замыкающих клеток в упругое состояние.
Общая площадь устьичных отверстий составляет лишь 1-2% площади листа. Несмотря на это, транспирация при открытых устьичных щелях достигает 50-70% испарения, равного по площади открытой водной поверхности.
Для соприкосновения листа с атмосферой имеются поры - устьица. Устьице - это отверстие (щель), ограниченная двумя замыкающими клетками. Устьица встречаются у всех наземных органов растения, но больше всего у листьев. Каждая замыкающая клетка устьица в отличие от клеток эпидермиса имеет хлоропласта. В них происходит фотосинтез, хотя с меньшей интенсивностью, чем в клетках мезофилла. Устьица - одно из оригинальных приспособлений, обладающих способностью открываться и закрываться в зависимости от насыщенности замыкающих клеток водой . Обычно устьичные отверстия ограничены двумя замыкающими клетками , стенки которых неравномерно утолщены. У двудольных растений замыкающие клетки бобовидной, или полулунной, формы, при этом их внутренние прилегающие друг к другу клеточные стенки более толстые, а внешние - более тонкие. Протопласты замыкающих клеток связаны в единое целое перфорациями в основании граничащих общих стенок. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на клеточные стенки, и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель.
В последнее время доказано, что для движения устьиц большое значение имеет также расположение микрофибрилл целлюлозы . Если обычно в клетках листьев целлюлозные фибриллы ориентированы в длину и в этом направлении утолщены, то в замыкающих клетках устьиц микрофибриллы организованы радиально, что усиливает устойчивость к процессу растяжения.
У злаков строение замыкающих клеток несколько иное. Они представлены двумя удлиненными клетками, на концах которых стенки более тонкие. При насыщении водой более тонкие стенки на концах растягиваются и раздвигают замыкающие клетки, благодаря чему образуется щель.
Число устьичных отверстий колеблется в зависимости от вида растений от 10 до 600 на 1 мм2 листа. У многих растений (75% видов), в том числе для большинства древесных, устьица расположены на нижней стороне листа. Диаметр устьичных щелей составляет всего 3-12 мкм. Устьица соединяют внутренние пространства листа с внешней средой. Вода поступает в лист через сеть жилок, в которых расположены сосудистые элементы. Возможны три пути испарения:
- через устьица - устьичная,
- кутикулу - кутикулярная,
- через чечевички - лентикулярная транспирация.
Впервые разграничение на кутикулярную и устьичную транспирацию было введено в 1877 г.
Основные типы устьичного аппарата листа растений.
- аномоцитный (у всех высших растений, кроме хвощей),
- диацитный (у папоротникови цветковых),
- парацитный (у папоротников, хвощей, цветковых и гнетовых),
- анизоцитный (только у цветковых),
- тетрацитный (главным образом у однодольных),
- энциклоцитный (у папоротников, голосеменныхи цветковых).
Устьица представляют собой высокоспециализированные образования эпидермы, состоящие из двух замыкающих клеток, между которыми имеется своеобразный межклетник, или устьичная щель.
Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Под щелью располагается дыхательная, или воздушная, полость, окруженная клетками мякоти листа. Клетки эпидермы, примыкающие к замыкающим, получили название побочных, или околоустьичных . Они участвуют в движении замыкающих клеток. Замыкающие и побочные клетки образуют устьичный аппарат .
Число и распределение устьиц на листе или побеге варьируют в зависимости от вида растений и условий жизни. Число их обычно колеблется от нескольких десятков до нескольких сотен на 1 кв.мм поверхности.
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при недостаточном водоснабжении в ночные часы, а иногда и днем тургор в замыкающих клетках понижается и щель замыкается, снижая тем самым уровень транспирации. С повышением тургора устьица открываются. Считают, что главная роль в этих изменениях принадлежит ионам калия. Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов. Первичный крахмал хлоропластов, превращаясь в сахар, повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и переходу замыкающих клеток в упругое состояние.
Общая площадь устьичных отверстий составляет лишь 1-2% площади листа. Несмотря на это, транспирация при открытых устьичных щелях достигает 50-70% испарения, равного по площади открытой водной поверхности.
Когда вы изучали строение отдельных органов и тканей, то в основном знакомились с теневыносливыми растениями, живущими в местах с достаточным увлажнением. Это наиболее типичная экологическая группа растений. Понимая общие закономерности строения и жизнедеятельности растений, зная характеристику этой экологической группы, вы сможете легче разобраться в особенностях организации других групп.
Светолюбивые растения . Свет оказывает большое влияние на форму растений. Растущие на открытом месте светолюбивые деревья, как правило, невысокие, разветвленные, с широкой кроной (сосна, береза, лиственница, белая акация). В лесу эти же деревья выглядят совершенно иначе, например сосна здесь имеет высокий, стройный ствол с кроной, расположенной на самой вершине [ 190]. А у сосны, выросшей на опушке, крона однобокая, годичные кольца древесины шире на освещенной стороне. Под влиянием затенения светолюбивые растения теряют боковые ветви.
Благодаря интенсивному освещению некоторые светолюбивые травянистые растения образуют низкорослые, часто розе- точные формы с укороченным стеблем (подорожник, одуванчик, горные растения), другие растения могут иметь достаточно высокий стебель (луговые травы, иван чай).
Светолюбивые растения имеют характерное строение листьев. Они обычно небольшие, плотные, с блестящей толстой кожицей и многочисленными устьицами. У многих растений листья покрыты восковым налетом или волосками, что предохраняет их от прямого воздействия солнечных лучей. У некоторых растений листья на стебле расположены вертикально (степные злаки) или повернуты ребром к падающим лучам солнца (эвкалипт). Это также избавляет лист от чрезмерного нагревания.
В клетках мякоти листа хлоропластов, как правило, немного, поэтому они имеют светло-зеленую окраску.
У светолюбивых растений хорошо развиты механические ткани и корневая система.
Тенелюбивые растения растут под пологом леса, в глубоких расщелинах и других местах, куда не проникают прямые лучи солнца (лишайник, вороний глаз, ветреница). В таких местах обычно повышенная влажность. Эти условия оказывают влияние на строение тенелюбивых растений.
Механические и проводящие ткани развиты слабо, поэтому побеги обычно хрупкие и нежные. Листовые пластинки у этих растений довольно крупные, тонкие. Кожица листа тонкая, ее клетки часто содержат хлоропласты. Устьица могут быть расположены на верхней и на нижней сторонах листа.
Хлоропласты в клетках листьев крупные, хлорофилла в них больше, чем у светолюбивых растений. Это обеспечивает возможность фотосинтеза при слабом, рассеянном освещении.
Растения водных и избыточно увлажненных мест обитания. Большинство живущих в воде растений имеет очень большую поверхность тела по отношению к его общей массе. Они поглощают воду и растворенные в ней вещества всей поверхностью тела, в связи с чем корневая система у них развита слабо, а иногда и совсем отсутствует (элодея, перистолистник). На погруженных в воду частях растений покровные ткани развиты слабо. Устьиц на подводных листьях нет.
У извлеченных из воды растений стебли и листья обвисают. Это связано с тем, что их механические ткани развиты очень слабо и растения могут сохранять вертикальное положение только в воде, которая их поддерживает.
Иное строение у листьев водных растений, плавающих на поверхности . На их верхней стороне, как и у сухопутных растений, образуются многочисленные устьица (на 1 мм 2 верхней стороны листа кувшинки их 460-500), обеспечивающие газообмен с воздушной средой и интенсивное испарение. Поверхность листа покрыта плотной глянцевой кожицей, она плохо смачивается, поэтому вода скатывается и не заливает устьица.
У всех водных растений сильно развита система межклетников, заполненных воздухом, а у некоторых образуется воздухоносная ткань.
Сухопутные растения, живущие в сильно увлажненной среде, совсем не выносят даже кратковременного иссушения. Это объясняется тем, что они плохо регулируют испарение, не могут сохранять в тканях необходимое количество воды и при засухе вянут и погибают. Среди этих растений можно выделить две группы. К первой относят растения, живущие в сильно увлажненных тенистых местах (в лесу это некоторые виды папоротников, кислица). Для растений этой группы характерны признаки тенелюбивых растений. У них обычно плохо развита корневая система, многочисленные устьица расположены на верхней и на нижней сторонах листа. У некоторых, кроме устьиц, есть особые образования - гидатоды (от греческих слов «гидор» - вода и «одос» - путь), через которые излишки воды удаляются из растений. Поэтому у многих растений, живущих при повышенной влажности почвы и воздуха, на листьях часто появляются капли воды. Это явление получило название «плач растений».
Ко второй группе относят растения, живущие в хорошо освещенных сырых местах, на переувлажненных почвах, во влажном воздухе, например калужница болотная, пушица, подмаренник болотный, рогоз.
Эти растения плохо переносят засуху, ветры. Корневая система у них обычно развита слабо, расположена поверхностно. Из-за недостатка воздуха во влажной почве в корнях и стеблях часто образуется система межклетников, развивается воздухоносная ткань. Некоторые особенности строения этих растений позволяют им быть более устойчивыми к действию прямых солнечных лучей. Кожица их листьев достаточно плотная, устьица расположены главным образом на нижней стороне. Механические ткани хорошо развиты.
Растения сухих мест обитания живут при значительном недостатке влаги, чаще всего в степях, полупустынях и пустынях. Растения этой экологической группы обычно обладают хорошо развитой корневой системой, многие из них запасают воду в тканях корня, стебля или листьев . Их листья имеют толстую, плотную кожицу, опушение или превращены в колючки (кактусы), устьиц немного, и расположены они, как у олеандра, - в углублениях . На листьях и других органах есть водонепроницаемый восковой налет (сахарный тростник), листья в период засухи могут быть свернуты в трубку (ковыль)
Таким образом, среди растений можно выделить несколько основных экологических групп: светолюбивые, теневыносливые, тенелюбивые, растения вод них а избыточно увлажненных мест обитания, растения мест достаточного увлажнения и растения сухих мест обитания. У растений каждой группы выработались приспособления, позволяющие им нормально расти и развиваться в определенных условиях обитания.
Хотя учёные давно знали об испарении воды поверхностью листа, первым, кто наблюдал устьица, был итальянский натуралист Марчелло Мальпиги , который это открытие опубликовал в 1675 году в своей работе Anatome plantarum . Однако он не понял их настоящую функцию. В то же время его современник Неемия Грю развил гипотезу об участии устьиц в вентиляции внутренней среды растения и сравнил их с трахеями насекомых . Прогресс в изучении наступил в XIX веке , и тогда же, в 1827 году , швейцарским ботаником Декандолем было впервые использовано слово „stoma“. Изучением устьиц в то время занимались Гуго фон Моль , который открыл основной принцип открывания устьиц и Симон Швенденер , классифицировавший устьица по типу их конструкции.
Некоторые аспекты функционирования устьиц продолжают интенсивно изучаться и в настоящее время; материалом в основном служат Коммелина обыкновенная (Commelina communis ), Боб садовый (Vicia faba ), Кукуруза сахарная (Zea mays ) .
Строение
Размеры устьица (длина) колеблются в пределах 0,01-0,06 мм (крупнее бывают устьица полиплоидных растений и у листьев, растущих в тени. Самые крупные устьица были обнаружены у вымершего растения Zosterophyllum , 0,12 мм (120 мкм) . Пора состоит из пары специализированных клеток, называемых замыкающими (cellulae claudentes ), которые регулируют степень открытости поры, между ними располагается устьичная щель (porus stomatalis ). Стенки замыкающих клеток утолщены неравномерно: направленные к щели (брюшные) толще стенок, направленных от щели (спинных). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель . Под щелью расположена подустьичная (воздушная) полость, окружённая клетками мякоти листа, через которую непосредственно и происходит газообмен . Воздух, содержащий диоксид углерода (углекислый газ) и кислород , проникает внутрь ткани листа через эти поры, и далее используется в процессе фотосинтеза и дыхании. Избыточный кислород, произведённый в процессе фотосинтеза внутренними клетками листа, выходит обратно в окружающую среду через эти же поры. Также, в процессе испарения через поры выделяются пары воды. Клетки эпидермиса , примыкающие к замыкающим, получили название сопровождающих (побочных, соседних, околоустьичных). Они участвуют в движении замыкающих клеток. Замыкающие и сопровождающие клетки образуют устьичный комплекс (устьичный аппарат). Наличие или отсутствие устьиц (видимые части устьиц называют устьичными линиями ) часто используют при классификации растений.
Типы устьиц
Число сопровождающих клеток и их расположение относительно устьичной щели позволяют выделить ряд типов устьиц:
- аномоцитный - сопровождающие клетки не отличаются от остальных клеток эпидермиса , тип весьма обычен для всех групп высших растений , за исключением хвойных ;
- диацитный - характеризуется только двумя сопровождающими клетками, общая стенка которых находится под прямым углом к замыкающим клеткам;
- парацитный - сопровождающие клетки располагаются параллельно замыкающим и устьичной щели;
- анизоцитный - замыкающие клетки окружены тремя сопровождающими, одна из которых заметно крупнее или мельче остальных, такой тип обнаружен только у цветковых растений;
- тетрацитный - четыре сопровождающие клетки, характерен для однодольных ;
- энциклоцитный - сопровождающие клетки образуют узкое колесо вокруг замыкающих клеток;
- актиноцитный - несколько сопровождающих клеток, радиально расходящихся от замыкающих клеток;
- перицитный - замыкающие клетки окружены одной побочной сопровождающей клеткой, устьице не соединено с сопровождающей клеткой антиклинальной клеточной стенкой;
- десмоцитный - замыкающие клетки окружены одной сопровождающей клеткой, устьице соединено с ней антиклинальной клеточной стенкой;
- полоцитный - замыкающие клетки окружены одной сопровождающей не полностью: к одному из устьичных полюсов примыкает одна или две эпидермальные клетки; устьице прикреплено к дистальной стороне единственной сопровождающей клетки, имеющей U-образную или подковообразную форму;
- стефаноцитный - устьице, окружённое четырьмя или более (обычно пять-семь) слабодифференцированными сопровождающими клетками, образующими более или менее отчётливую розетку;
- латероцитный - такой тип устьичного аппарата рассматривается большинством ботаников как простая модификация аномоцитного типа.
Расположение устьиц
Двудольные растения, как правило, в нижней части листа имеют больше устьиц, чем в верхней. Это объясняется тем, что верхняя часть горизонтально-расположенного листа, как правило, лучше освещена, и меньшее количество устьиц в ней препятствует избыточному испарению воды. Листья с устьицами, расположенными на нижней стороне, называются гипостоматическими.
У однодольных растений наличие устьиц в верхней и нижней части листа различно. Очень часто листья однодольных растений расположены вертикально, и в этом случае количество устьиц на обоих частях листа может быть одинаково. Такие листья называются амфистоматическими.
У плавающих листьев на нижней части листа устьица отсутствуют, так как они могут впитывать воду через кутикулу . Листья с устьицами, расположенными на верхней стороне, называются эпистоматическими. У подводных листьев устьица отсутствуют совсем.
Устьица хвойных растений обычно спрятаны глубоко под эндодермой, что позволяет сильно снизить расход воды зимой на испарение, а летом - во время засухи.
У мхов (исключение антоцеротовые) настоящие устьица отсутствуют.
Устьица также различаются по уровню расположения относительно поверхности эпидермиса. Некоторые из них расположены вровень с другими эпидермальными клетками, другие подняты выше или погружены ниже поверхности. У однодольных, листья которых растут преимущественно в длину, устьица образуют правильные параллельные ряды, тогда как у двудольных они располагаются беспорядочно.
Углекислый газ
Так как углекислый газ является одним из ключевых реагентов в процессе фотосинтеза, у большинства растений устьица в дневное время открыты. Проблема состоит в том, что при входе воздух смешивается с парами воды, испаряющимися из листа, и поэтому растение не может получить углекислый газ, одновременно не потеряв некоторое количество воды. У многих растений существует защита от испарения воды в виде закупоривающих устьица восковых отложений.
История исследований
Хотя учёные давно знали об испарении воды поверхностью листа, первым, кто наблюдал устьица, был итальянский натуралист Марчелло Мальпиги , который это открытие опубликовал в 1675 году в своей работе Anatome plantarum . Однако он не понял их настоящую функцию. В то же время его современник Неемия Грю развил гипотезу об участии устьиц в вентиляции внутренней среды растения и сравнил их с трахеями насекомых . Прогресс в изучении наступил в XIX веке , и тогда же, в 1827 году , швейцарским ботаником Декандолем было впервые использовано слово „stoma“. Изучением устьиц в то время занимались Гуго фон Моль , который открыл основной принцип открывания устьиц и Симон Швенденер , классифицировавший устьица по типу их конструкции.
Некоторые аспекты функционирования устьиц продолжают интенсивно изучаться и в настоящее время; материалом в основном служат Коммелина обыкновенная (Commelina communis ), Боб садовый (Vicia faba ), Кукуруза сахарная (Zea mays ).
Строение
Размеры устьица (длина) колеблются в пределах 0,01–0,06 мм (крупнее бывают устьица полиплоидных растений и у листьев, растущих в тени. Самые крупные устьица были обнаружены у вымершего растения Zosterophyllum , 0,12 mm (120 µm) Пора состоит из пары специализированных клеток, называемых замыкающими (cellulae claudentes) , которые регулируют степень открытости поры, между ними располагается устьичная щель (porus stomatalis) . Стенки замыкающих клеток утолщены неравномерно: направленные к щели (брюшные) толще стенок, направленных от щели (спинных). Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель . Под щелью расположена подустьичная (воздушная) полость, окружённая клетками мякоти листа, через которую непосредственно и происходит газообмен . Воздух, содержащий диоксид углерода (углекислый газ) и кислород , проникает внутрь ткани листа через эти поры, и далее используется в процессе фотосинтеза и дыхании. Избыточный кислород, произведённый в процессе фотосинтеза внутренними клетками листа, выходит обратно в окружающую среду через эти же поры. Также, в процессе испарения через поры выделяются пары воды. Клетки эпидермиса, примыкающие к замыкающим, получили название сопровождающих (побочных, соседних, околоустьичных). Они участвуют в движении замыкающих клеток. Замыкающие и сопровождающие клетки образуют устьичный комплекс (устьичный аппарат). Наличие или отсутствие устьиц (видимые части устьиц называют устьичными линиями ) часто используют при классификации растений.
Типы устьиц
Число сопровождающих клеток и их расположение относительно устьичной щели позволяют выделить ряд типов устьиц:
- аномоцитный - сопровождающие клетки не отличаются от остальных клеток эпидермиса , тип весьма обычен для всех групп высших растений , за исключением хвойных ;
- диацитный - характеризуется только двумя сопровождающими клетками, общая стенка которых находится под прямым углом к замыкающим клеткам;
- парацитный - сопровождающие клетки располагаются параллельно замыкающим и устьичной щели;
- анизоцитный - замыкающие клетки окружены тремя сопровождающими, одна из которых заметно крупнее или мельче остальных, такой тип обнаружен только у цветковых растений;
- тетрацитный - четыре сопровождающие клетки, характерен для однодольных ;
- энциклоцитный - сопровождающие клетки образуют узкое колесо вокруг замыкающих клеток;
- актиноцитный - несколько сопровождающих клеток, радиально расходящихся от замыкающих клеток;
- перицитный - замыкающие клетки окружены одной побочной сопровождающей клеткой, устьице не соединено с сопровождающей клеткой антиклинальной клеточной стенкой;
- десмоцитный - замыкающие клетки окружены одной сопровождающей клеткой, устьице соединено с ней антиклинальной клеточной стенкой;
- полоцитный - замыкающие клетки окружены одной сопровождающей не полностью: к одному из устьичных полюсов примыкает одна или две эпидермальные клетки; устьице прикреплено к дистальной стороне единственной сопровождающей клетки, имеющей U-образную или подковообразную форму;
- стефаноцитный - устьице, окружённое четырьмя или более (обычно пять-семь) слабодифференцированными сопровождающими клетками, образующими более или менее отчётливую розетку;
- латероцитный - такой тип устьичного аппарата рассматривается большинством ботаников как простая модификация аномоцитного типа.
У двудольных распространённым является парацитный тип устьиц. Замыкающие клетки почковидной (бобовидной) формы - такими они видны с поверхности листа - несут хлоропласты , тонкие неутолщённые участки оболочки образуют выступы (носики) закрывающие устьичную щель.
Наружные стенки замыкающих клеток обычно имеют выросты, что хорошо видно на поперечном разрезе устьица. Пространство, ограниченное этими выростами, называют передним двориком. Нередко аналогичные выросты наблюдаются и у внутренних оболочек замыкающих клеток. Они образуют задний дворик, или внутренний, соединённый с крупным межклетником - подустьичной полостью.
У однодольных парацитное строение устьиц отмечено у злаковых . Замыкающие клетки имеют гантелевидную форму - сужены в средней части и расширены на обоих концах, при этом стенки расширенных участков очень тонкие, а в средней части замыкающих клеток сильно утолщены. Хлоропласты располагаются в пузыревидных окончаниях клеток.
Движение замыкающих клеток
Механизм движения замыкающих клеток весьма сложен и неодинаков у разных видов. У большинства растений при неодинаковом водоснабжении в ночные часы, а иногда и днём тургор в замыкающих клетках понижается, и устьичная щель замыкается, снижая тем самым уровень транспирации . С повышением тургора устьица открываются. Считают, что главная роль в изменении тургора принадлежит ионам калия . Существенное значение в регуляции тургора имеет присутствие в замыкающих клетках хлоропластов. Первичный крахмал хлоропластов, превращаясь в сахар , повышает концентрацию клеточного сока. Это способствует притоку воды из соседних клеток и повышению тургорного давления в замыкающих клетках.
Расположение устьиц
Двудольные растения, как правило, в нижней части листа имеют больше устьиц, чем в верхней. Это объясняется тем, что верхняя часть горизонтально-расположенного листа, как правило, лучше освещена, и меньшее количество устьиц в ней препятствует избыточному испарению воды. Листья с устьицами, расположенными на нижней стороне, называются гипостоматическими.
У однодольных растений наличие устьиц в верхней и нижней части листа различно. Очень часто листья однодольных растений расположены вертикально, и в этом случае количество устьиц на обоих частях листа может быть одинаково. Такие листья называются амфистоматическими.
У плавающих листьев на нижней части листа устьица отсутствуют, так как они могут впитывать воду через кутикулу . Листья с устьицами, расположенными на верхней стороне, называются эпистоматическими. У подводных листьев устьица отсутствуют совсем.
Устьица хвойных растений обычно спрятаны глубоко под эндодермой, что позволяет сильно снизить расход воды зимой на испарение, а летом - во время засухи.
У мхов (исключение антоцеротовые)настоящие устьица отсутствуют.
Устьица также различаются по уровню расположения относительно поверхности эпидермиса. Некоторые из них расположены вровень с другими эпидермальными клетками, другие подняты выше или погружены ниже поверхности. У однодольных, листья которых растут преимущественно в длину, устьица образуют правильные параллельные ряды, тогда как у двудольных они располагаются беспорядочно.
Углекислый газ
Так как углекислый газ является одним из ключевых реагентов в процессе фотосинтеза, у большинства растений устьица в дневное время открыты. Проблема состоит в том, что при входе воздух смешивается с парами воды, испаряющимися из листа, и поэтому растение не может получить углекислый газ, одновременно не потеряв некоторое количество воды. У многих растений существует защита от испарения воды в виде закупоривающих устьица восковых отложений.
Примечания
Литература
- Атлас по анатомии растений: Учеб. пособие для вузов / Бавтуто Г. А., Ерёмин В. М., Жигар М. П.. - Мн. : Ураждай, 2001. - 146 с. - (Учеб. и учеб. пособия для вузов). - ISBN 985-04-0317-9
- [[:en:{{{1}}}|]] Шаблон:Цитирование монографии
Сноски
Wikimedia Foundation . 2010 .
Статьи по теме